Virulent effect of solar radiation at the boundary between the UVB and UVA ranges
Virulent effect of solar radiation at the boundary between the UVB and UVA ranges

Abstract
The results of an analysis of the frequency of viral pandemics from 2008 to 2019 are presented, including statistics on Ebola haemorrhagic fever, influenza AH1N1, AH7N9 influenza, MERS-CoV coronavirus, and HIV-1 infected patients in the Russian Federation from 1996 to 2019 (23–24 solar activity cycles) with solar radiation intensity at a wavelength of ~λ315 nm, representing the boundary between the UVB (280–315 nm) and UVA (315–400 nm) ranges, as well as the boundary of the relative spectral bactericidal efficiency curve (205–315 nm). A close relationship has been established between viral pandemics and HIV-infected patients (a total of ~ 100,000 people) with a change in intensity ~λ315 nm (r ~ 0.85) in solar activity cycles. A conclusion has been made about the virulent effect of solar radiation ~ 315 nm on the development of viral pandemics.
1. Введение
Посвящается труду А.Л. Чижевского
«Эпидемические катастрофы и периодическая деятельность Солнца» (1930 г.)
Изменения спектральной солнечной светимости на участке приходящего к земной поверхности излучения UVB (297–315 nm) — UVA (315–400 nm) обусловлены глобальными солнечными механизмами с временным диапазоном от минут до десятилетий. К ним относятся:
1) солнечные осцилляции, воздействующие на UVB-UVA с частотой «пятиминутных колебаний»
;2) вращение солнечной атмосферы (~ 27 сут);
3) 11-летний цикл солнечной активности (СА), с которым тесно коррелирует излучение UVB .
В работе , посвященной анализу связи глобальных эпидемий (Global Epidemics, GE), вызванных вирусом лихорадки Эбола, гриппа AH1N1, AH7N9 и коронавируса MERS-CоV, с изменением солнечной активности (СА) в 24 цикле, получена корреляция динамики GE с излучением UV (r~0,83) на границе кривой относительной спектральной бактерицидной эффективности λ315 nm, представляющая также границу диапазонов приходящего к земной поверхности солнечного ультрафиолета UVB (280–315 nm) и UVA (315–400 nm). На рис. 1 представлена гистограмма интенсивности ~ λ315 nm и годовых значений GE, за период с 2008 г. по 2019 г. (24 цикл СА) по данным с сайтов Umweltphysik IUP
и World Health Organization . Значения GE на левой оси ординат рис. 1 представляют суммарную частоту вспышек высокопатогенных вирусов геморрагической лихорадки Эбола, гриппа AH1N1, AH7N9 и коронавируса MERS-CоV (родственный SARS-CoV-2).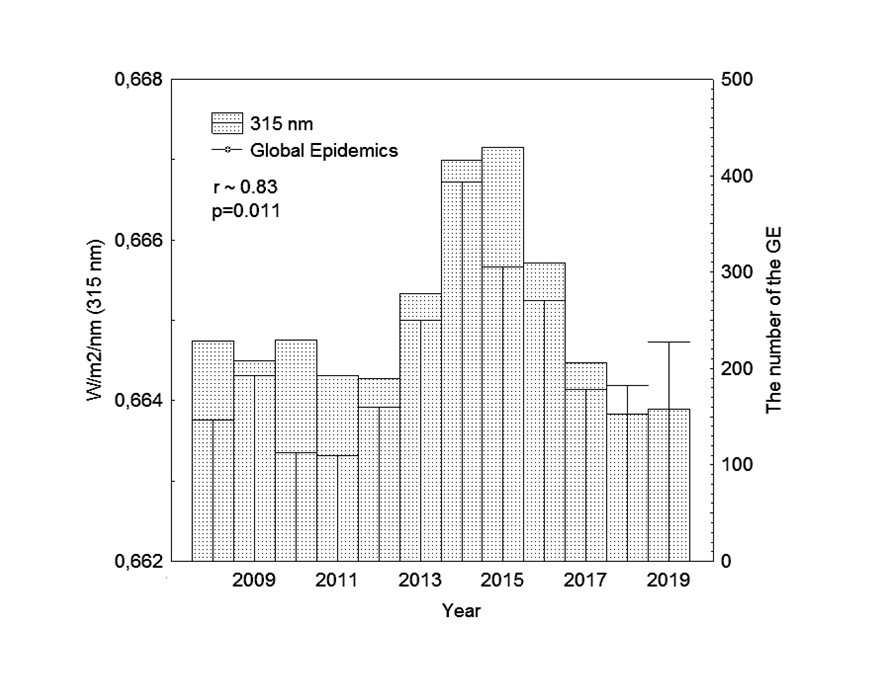
Рисунок 1 - Гистограмма случаев вирусных пандемий (Global Epidemics, GE) и интенсивности солнечного излучения на границе диапазонов UVB-UVA (λ315 nm) с 2008 г. по 2019 г. (24 цикл СА)
Для подтверждения выводов о связи различных групп вирусов с излучением UV в 24 цикле СА мы использовали данные по численности ВИЧ-инфицированных в Российской Федерации за период с 1997 г. по 2019 гг. (https://www.crie.ru/). Данные по интенсивности λ315nm выбирались на сайте https://lasp.colorado.edu/. В качестве показателя СА выбран композитный индекс MgII (280 nm), характеризующий глобальное изменение UV на Солнце в 11-летнем цикле СА (https://www.iup.uni-bremen.de/deu). Статистика вируса иммунодефицита человека ВИЧ-1 (HIV-1) в значение суммарной частоты не применялась, т.к. ВИЧ-1 отличается от вышеупомянутых вирусов повышенной мутагенностью, скоростью размножения, поражением клеток «первой линии защиты» иммунной системы (лимфоциты Т-хелперы-1, макрофаги и нейтрофилы), последующей иммуносупрессией и активацией ранее «спящих» патогенов. Последнее может привести к гибели организма человека, как итогу медленно развивающегося комплексного заболевания СПИД. Описанные особенности HIV-1 предполагают существование как специфических, так и общих с другими вирусами механизмов влияния интенсивности излучения границы UVB-UVA на вирулентность HIV-1.
2. Результаты
Использовались данные по численности HIV-1 инфицированных пациентов в РФ за период с 1997 г. по 2019 г. (всего ~ 100000 человек). Данные выбирались на сайтах www.hivrussia.info/dannye-po-vich-infektsii-v-rossii и www.lasp.colorado.edu. В качестве показателя СА выбран композитный индекс MgII (280 nm), характеризующий изменение интегрального UV в солнечной атмосфере . В итоге обработки данных на рис. 2а представлено распределение численности HIV-1 инфицированных (по годовым значениям) с линейным трендом, обусловленным ростом парентеральных контактов при употреблении наркотиков, и без тренда. С последним сопоставляли ход интенсивности λ315 nm (рис. 2b) и композитного индекса Mg ll (рис. 2с). На рис. 3 представлено аналогичное рис. 2 сопоставление HIV-1 и частоты вирусных пандемий (GE) на фоне излучения λ315 nm (23–24 циклы СА). Пунктиром GE обозначено вероятное изменение GE в 23 цикле СА, т.к. статистика по GE за данный период в архиве WHO не найдена . Как видно на рисунке графики демонстрируют идентичность хода годовых значений GE и HIV-1.
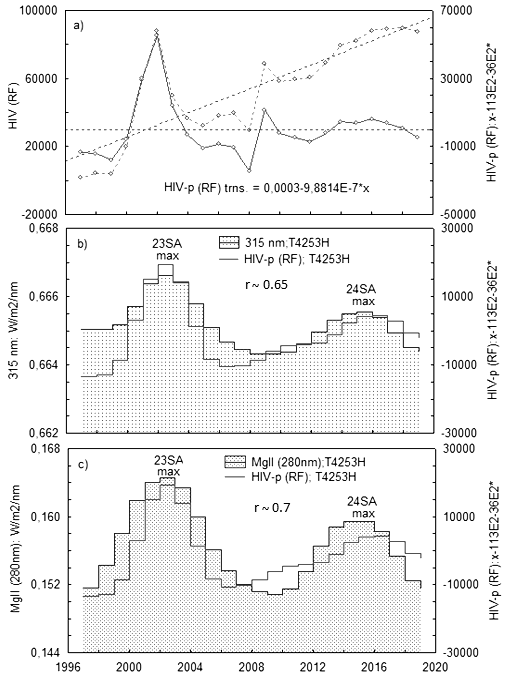
Рисунок 2 - Временной ряд HIV-1-инфицированных пациентов в РФ с линейным трендом, обусловленным ростом парентеральных контактов при употреблении наркотиков, и без тренда (a); сопоставление временного хода HIV-1 с излучением ~λ315 nm в интервале 23–24 циклов СА (b); сопоставление хода HIV-1 с композитным индексом Mg II (280 nm) в интервале 23–24 циклов СА (с)
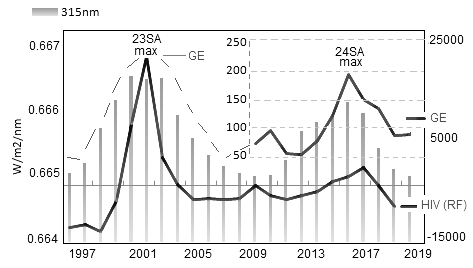
Рисунок 3 - Сравнение численности HIV-1-инфицированных в РФ и частоты планетарных пандемий (GE) на фоне излучения λ315 nm за период 1997–2019 гг. (23–24 циклы СА)
3. Энергия диапазонов UVB-UVA в фотобиологических процессах
Механизм бактерицидного эффекта UV в отношении разных вирусов заключается в повреждении его нуклеиновой кислоты, что блокирует размножение. Однако энергия квантов UVB-UVA способна запускать в клетках «хозяина» несколько типов сходных «обходных» путей, облегчающих размножение вируса и, одновременно, снижающих иммунорезистентность организма:
1. UVВ и цитокины. Известно, что медиаторами межклеточного взаимодействия в иммунной системе человека являются про- и противовоспалительные цитокины, а также EGF — эпидермальный фактор роста . Например, после облучения UVB кератиноциты эпидермиса секретируют цитокин IL-10 (Interleukin-10), подавляющий антиген-презентацию клетками Лангерганса, — иммунными клетками кожи. Кроме того, UVB подавляет секрецию лимфоцитами (Т-хелперами-1) ряда про-воспалительных цитокинов: IL-1,-6,-12 и фактора некроза опухоли альфа (TNFa)
, . Последнее, совместно с пролиферативным эффектом EGF, создает условия для онкогенеза.2. UVA и Митохондрии (МТХ). В МТХ кератиноцитов и фибробластов кожи, а также в МТХ Т-хелперов-1 UVA способствует образованию синглетного кислорода, супероксид аниона и АКМ. Они вызывают перекисное окисление липидов и трансформацию белков мембран иммунокомпетентных клеток (мишеней для HIV-1) с их последующим апоптозом , чем облегчают подавление иммунного ответа этим вирусом.
3. UVB-UVA, МТХ и Translocator protein (TSPO). Белок TSPO 18 kDa, поглощая фотон UV, транспортирует холестерин между мембранами МТХ и обеспечивает его взаимодействие со стероидогенным фактором StF-1 и последующий синтез стероидных гормонов в клетках стероидогенных тканей ,
, . Ключевая роль TSPO в стероидогенезе обусловливает его участие в стероид-зависимом контроле иммунного ответа, стресс-адаптации, клеточной пролиферации и апоптозе.В а,б,с- путях ключевой структурой являются МТХ — «генератор энергии» клетки, что важно для взаимодействия разных путей опосредования эффектов UV (внеклеточного источника энергии) и адаптивного варьирования их одновременного или последовательного действия. Например, энергетическая «цена» полимеризации TSPO при λ254 nm требует участия АКМ
, , тогда как при λ302 nm такой необходимости нет. Далее, интенсивность λ302 nm в 120–360 J/m2 вызывает полимеризацию ТSРО и усиленный стероидогенез, при увеличении интенсивности до 480 J/m2 той же длины волны происходит деполимеризация белка и снижение его стероидогенной функции . Описанные особенности изменения структуры и функций UV-чувствительного белка TSPO в клетках животных и человека объясняют полученные в данной работе факты зависимости числа инфицированных HIV-1 и числа вирусных пандемий от интенсивности границы диапазонов UVB-UVA (~ λ315 nm) в 11-летнеем цикле СА. Если фоточувствительные процессы в МТХ усиливают ацидоз, создавая условия для проникновения вируса в клетки-мишени и его размножения, то мишени UV в цитоплазме и ядре реализуют и другие эффекты, например, для витаминов Д3, В2 (рибофлавина), а также криптохромов:1. UVB и витамин D3. Синтез витамина 1,25(ОН) D3 начинается в кератиноцитах кожи под влиянием UVB
. Рецептор витамина D3 является транскрипционным фактором, действующим совместно с рецептором гормонов щитовидной железы — три- и тетра-йодтиронинов (усилители синтеза АТФ в МТХ). Витамин D3 необходим для усвоения кальция и фосфора в костной ткани и Са-зависимой активации иммунокомпетентных клеток. Поскольку Са2+ при ацидозе играет роль буфера, то витамин D3 противодействует эффектам HIV-1.2. UVB и FAD-содержащие белки цитоплазмы. Фотоны излучений диапазонов UVB-UVA в клетке-мишени для вируса могут активировать фоточувствительные белки, содержащие последовательность флавинаденилдинуклеотид (FAD, ко-фактор многих окислительно-восстановительных реакций). У животных и человека наиболее известны криптохромы CRY1/CRY2 и витамин В2 или рибофлавин, содержащий FAD в комплексе с флавинмононуклеотидом (FMN). Криптохромы CRY1-CRY2 у животных и растений входят в состав молекулярного внутриклеточного механизма поддержания циркадианного (околосуточного) ритма жизнедеятельности. При активации фотоном FAD в составе CRY1 он с помощью своего неспаренного электрона последовательно взаимодействует с цепочкой остатков аминокислоты триптофана в молекуле CRY, каждый раз образуя пару радикалов FAD-Trp, обладающую магнито-чувствительностью. Так, под влиянием фотонов UV белок приобретает функции фото- и магнито-сенсора
, а также становится первопричиной появления цепей свободных радикалов и нарастания в клетке ацидоза, облегчая размножение вируса. В составе рибофлавина FAD, активированный фотоном UVB, также может быть причиной роста концентрации свободных радикалов, разрушающих структуры внеклеточных вирусов и тем самым угнетающих их и блокирующих распространение патогена. Это может быть универсальным механизмом угнетающего действия рибофлавина, под воздействием света в отношении вирусов, микробов, грибов и паразитов . Вместе с тем, рибофлавин активирует иммунную защиту организма, обусловливая совместно с криптохромами суточные и сезонные ритмы активности иммунной системы. Благодаря вариациям UVB-UVB в 11-летнем цикле СА, инфрадианные многолетние макроритмы активности иммунной защиты и вирусных пандемий могут быть прогнозируемы . В свою очередь, усиление интенсивности UVB-UVA в ходе цикла СА является необходимым условием для формирования вирусных пандемий.4. Заключение
В целостной картине воздействия Солнца на патогенную среду излучение UVB играет роль фотобиологического фактора. Основным источником энергии для вирусов являются окислительно-восстановительные реакции клеток организма хозяина, опосредующих действие UVB-UVA. Вирусы не обладают способностью к продукции энергии, однако вопрос об использовании ими энергии СА и наличии в их структуре фоточувствительных белков остаётся открытым (в геноме вируса обнаружен ген родопсина, но не белок).
Обобщая рис. 3 следует вывод о высокой чувствительности различных вирусов к интенсивности на длине волны ~ λ315 nm, определяющей условие для начальной стадии интенсификации (или редукции) патогенной среды.
Если рассматривать «вирус и окружающие его клетки хозяина» как единую двухкомпонентную систему, опосредующую влияние UVB, то можно проследить черты её адаптации к солнечным факторам. Они проявляются, например, в появлении свойств магнито-сенсора у белка криптохрома (CRY1) после облучения UVB. Другой пример — бистабильность UV-воспринимающих белков человека CRY1 и опсина 5 (OPN5), проявляющаяся в сочетании одновременной или последовательно сменяющейся чувствительности к излучению оптического диапазона солнечного спектра . При этом UV-чувствительные белки выступают в роли фотосенсибилизатора, который вызывает возникновение ионизированных форм кислорода , свободных радикалов и в основном усиливает действия патогена. «Совмещение» свойств фото- и магнитосенсоров у ключевых для метаболизма белков, является, по-видимому, также одной из адаптаций живых организмов к воздействию излучения UVB на границе ~ λ315 nm. Не исключено, что реакция вирусов на излучение различных длин волн в диапазоне UVB (280–315 nm) может усиливаться/уменьшаться за счет региональных особенностей орографии и атмосферы.